
Types of Songs and Calls/Sonograms
Brennan and Jones (2016) conducted an observational study, where they recorded 3,088 songs from 109 different male Veeries in four regions across the Appalachian Mountains. They differentiated songs based on structural elements such as syllables and phrases, revealing song repertoire and patterns in song delivery. The song repertoire was found to be 1-6 songs when basing the song on structure alone, however, increased to 3-7 songs when focusing on both structure and frequency. Two syllables made up the entirety of the songs that the Veery sings,labeled B and C. There is an introductory syllable, but it starts off each song and so isn’t a defining feature. The arrangement of these two syllables is what differs one song from another. Figure 1 is a spectrogram of a song that the Veery sings and it reveals how only two syllables are used in the song, and that two B syllables make up a phrase.The song consists of two phrases, each composed of two identical syllables. Four song types were found to be the most common where 88% of birds had a BBCC song in their repertoire, 8% had a BBB and
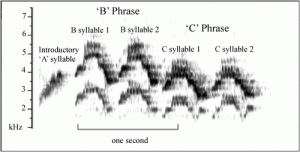
Fig. 1 Spectrogram of “BBCC” song (Brennan & Jones, 2016)
BBC arrangement, and lastly 6% had a BCC song. The rest of the songs recorded were isolated in certain locations and comprised such a small percentage of the whole population that they were negligible to the study. A recording of the Veery song can be found here:Veery song with spectrogram. Figure 2 reveals the oscillating peak frequencies of the same song for an individual, meaning while each song is structurally similar, it is sang at different frequencies. This figure illustrates how
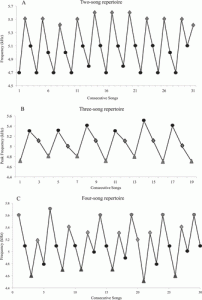
Fig 2. Oscillating peak frequencies of song across 3 song repertoires (Brennan & Jones, 2016)
oscillating peak frequency patterns are observed in both birds with small and large song repertoires. Analyzing song from more than just a structural perspective is critical as it in fact increased the song repertoire of the Veery to 3-7 songs.
The role of song in this thrush species is delayed until after bond formation and is most frequent during female incubation. (Heckscher, 2007). Dilger (1956) confirmed that males rarely sing before bond formation, and found that song was used as a hostile vocalization in the nest when the female was incubating. Song therefore doesn’t play in role in courtship or defining new territories. Song can also be said to not be a sexually selected trait in this species as a female chooses a male without having heard his song.
Heckscher’s (2007) six-year observational study concluded that the Veery has 6 distinct types of calls. The majority of the calls were single, modulating frequency notes. The first is characterized as having an initial rise in frequency, before sweeping downward from a high to low frequency (Fig. 3). This note sounds harsh due its endpoint having a broadband frequency. The second call is another downward sweeping note from high to low frequency, however, is tonal because of its continuous narrowband frequency (Fig. 4). There was a strong correlation between harsh/tonal sounds and communication distance. Harsh sounds give away the location of the signaler, and as a result are only used in long distance communication. Tonal calls on the other hand reveal imprecise location, making it safe to signal in short distances. Due to the dense understory that the Veery inhabits, vision is impaired, so the Veery relies on a communication system that either reveals or conceals location. The next four calls are each structurally dissimilar to the first two down sweeping calls. They are a high frequency, narrowband whistle(Fig.5), a narrowband, downward call (Fig. 6), a u-shaped call that is restricted within a very limited frequency range (Fig. 7), and lastly a chatter that appears on a spectrogram as a clutter of sounds having broadband and varying frequencies (Fig. 8). Chatter is primarily used in the beginning of the breeding season as a tool defining male territories and is also used by both males and females in aggressive encounters. U-shaped calls were primarily used during fledgling management and were absent in defining territory. The downward call, primarily expressed my males, is a neutral signal, neither revealing location nor having any aggressive motives. Lastly, the whistle is used in the beginning of the breeding season in courtship. This complex communication system evolved as a result of the environment the Veery resides in. The dense understory that this thrush species inhabits impairs vision, and as a result complex calls revealing location, spatial awareness, and aggression are necessary for survival.

Fig. 3 Harsh call (Heckscher, 2007)

Fig. 4 Tonal call (Heckscher, 2007)

FIg. 5 Whistle (Heckscher, 2007)

FIg. 6 Downward call (Heckscher, 2007).

Fig. 7 U-shaped call (Heckscher, 2007)

FIg. 8 Chatter call (Heckscher, 2007)
LEARNING
There is a lack of research on the song development of the Veery, however there is consensus that their song is learned. Unlike suboscine birds that innately produce their species’ song, the Veery is an oscine bird, which learns its species’ song through a song learning process guided by an innate song and social interactions that results in the crystallized song (Gill, 2007). Song development of the Veery begins with the sensory phase, where young Veeries listen to a tutor’s song and commit it to memory. After they have committed the tutor’s song to memory, the young Veery enters the motor or production phase. During this phase, Veeries practice and rehearse their memorized song in order to crystallize it (Bell et al, 1998). The learning period for the Veery is still unknown and there is debate over if the Veery learns its songs within the first summer during natal dispersal similar to the White-crowned sparrow or if it is similar to that of young Chaffinch males who can continue song learning up to the next breeding season (Marova 2010). Although Veeries do learn their song from a tutor, they are guided through the song development process by an innate song. A study conducted in 2013, found that 88% of the studied Veeries had the ‘BBCC’ song in their repertoire and that the other main songs recorded were variations of this one template song (Brennan, 2013). This widespread frequency of the ‘BBCC’ song is because it is a genetically innate template song of the Veery, therefore there is an inborn tendency to sing this specific song (Brennan, 2013).
DIALECT
There is some evidence of the Veery having multiple dialects. During Brennan’s study in 2015, he found that there were differences along the Appalachian Mountains in the frequency ranges of the Veery song. During the study, Brennan recorded the songs of male Veeries from states along the Appalachian and found that there was a distinctive difference between the songs of the North Carolina Veery, in some cases the West Virginia Veery, and the Veeries from the other states. In this case the southern Veeries had a lower frequency when singing their song while the northern Veeries had a higher frequency (Brennan, 2013). The distinct frequency ranges sung by southern Veeries and northern Veeries are possibly due to the restricted gene flow between the northern and southern populations (Brennan, 2013). These two types of Veeries makeup half of the the total subspecies of the Veery. The four subspecies of the Veery are C. f. salicicola, C. f. subpallidus, C. f. fuscescens, and C. f. fuliginosus (Collar et al, 2018). There has been little research on the subspecies, but the area in which they are found is known; the C. f. salicicola is found in southwest Canada and western USA, the C. f. subpallidus is found in northwestern USA, the C. f. fuscescens is found in southeast Canada and eastern USA, and the . f. fuliginosus is found in east Canada (Collar et al, 2018). In order to fully understand the subspecies of the Veery and to see if these subspecies are diverging, there needs to be more research concerning gene flow variation and song development.
References
Bell, D. A., Trail, P. W., & Baptista, L. F. (1998). Song learning and vocal tradition in Nuttalls white-crowned sparrows. Animal Behaviour, 55(4), 939-956.
Brennan, C. (2013) “Singing Behavior and Geographic Variation in the Songs of the Veery (Catharus Fuscescens) Across the Appalachian Mountains”. ETD Archive. Paper 511.
Brennan, C. & Jones, A. (2016). Song structure and cadence of the Veery (Catharus fuscescens) in the Appalachian Mountains. Wilson Journal of Ornithology. 128(1),75-85.
Collar, N., Christie, D.A., de Juana, E. & Kirwan, G.M. (2018). Veery (Catharus fuscescens). In: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A. & de Juana, E. (eds.). Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona.
Dilger, W. (1956). Hostile behavior and reproductive isolating mechanisms in the avian genera Catharus and Hylocichla. 73(3), 313-353.
Gill, F. B. (2007). Ornithology, Third Edition. W. H. Freeman and Company, New York, USA.
Heckscher, C. (2007). Use of the Veery (Catharus fuscescens) call repertoire in vocal communication. Ph.D. dissertation, University of Delaware, Newark, DE.
Marova, I. M., V. V. Ivanitskii, and O.D. Veprintseva. (2010). Individual, population, and geographic differentiation in advertising song of the Blyth’s Reed Warbler, Acrocephalus dumetorum (Sylvidae). Biology Bulletin 37(8),846–860.
