
The Bobolink (Dolichonyx oryzivorus)-Song
BOBOLINKS AND SONG
Bobolinks have two variants of primary song, an alpha variant and a shorter beta variant. Their primary song has been described by Bent (1958) as a song that even the mockingbird cannot reproduce. It is a “bubbling delirium of ecstatic music that flows from the gifted throat of the bird like sparkling champagne”. Admittedly, it is pretty difficult to describe the song of the Bobolink just by listening to it. Wittenberger (1983) ventured to phonetically describe the dialect of southeastern Oregon Bobolinks. To him, “the alpha song sounds something like ‘puck puck pi, deedla eh ah, eeee-ew, d-t-d-t dee,’” followed by “rapidly warbled gurgling noises (of variable duration), ‘ew.’” and “the beta song sounds something like ‘pete-n pete-n d-t-d-t dee,’” also followed by “rapidly warbled gurgling noises (of variable duration), ‘ew.’”.

Sound spectrographs of the alpha (top) and beta (bottom) song types by male Bobolinks in Oregon. Taken from Wittenberger (1983).
Bobolinks possess an extremely localized song dialect. Songs may be shared with birds within a very concentrated geographical area, but outside of a certain range, the songs may differ completely. Avery (1977) performed a study on Bobolinks in North Dakota and Minnesota and found that little similarity was drawn between populations unless they were less than one kilometer apart.
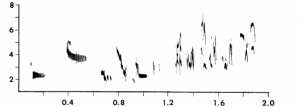
Sound spectrogram of male Bobolink song from Chase Lake in Minnesota. See how different it is from the Oregon Bobolink from above! Taken from Avery (1977).
To explain this localized dialect, Avery (1977) theorized that a male Bobolink probably acquires his songs during his first breeding season by copying the songs of an older male. Therefore, a Bobolink will most likely learn its song from its father. However, no studies have been done to show this is actually the case.
The following is a sample of male Bobolink song from West Virginia:
The beta song of the Bobolink has been previously thought by Wittenberger (1983) to carry a more aggressive message than the alpha song. The alpha song, meanwhile, was thought to be involved with mate attraction. However, experiments performed by Capp (1992), using variable seasonal effects, location effects, and playback experiments have demonstrated that there is no evidence that the two different song type convey different messages. Capp instead found evidence to support that the beta and alpha songs are used in a form of song matching, in which the beta song is sung in response to the alpha song and vice versa.
It is interesting to note that despite this, Ammer (1999) has found that male Bobolinks include more song variants when in the presence of a potential breeding partner. Additionally, he concluded from experimental data that song versatility is significantly greater in an intersexual context rather than an intrasexual context.
Bobolink song may, in the future, be influenced by nearby noise and structures. A study done on chipping sparrows, another grassland bird, has found that noise, urban structures, and vegetation have each influenced the structure of chipping sparrow song (Job 2016). The closer urban structures were, the more decreased the peak frequency was. Vocal deviation was also found to become increasingly larger and more negative with the increasing presence of urban structures and noise, respectively. It may be a concern that the ambient noise from the structures themselves may inadvertently affect the study, but Koper (2016) showed that the effects are negligible within a grassland environment.
WHERE SHOULD FUTURE RESEARCH FOCUS?
There has been a number of studies done on figuring out whether the alpha and beta song variants possess different meanings, but no studies, as far as that was available to us, that detailed how the Bobolink learns its song.
Therefore, a controlled experiment should be done to determine exactly how the Bobolink learns its song. Bobolink chicks should be raised in isolation and divided into three groups, one control and two experimental. The control would be raised with Bobolink song from its biological parents. One experimental group is exposed to Bobolink song from a different dialect, while the other is exposed to song from a different bird species.
If the chicks in the first experimental group copy the dialect, and the chicks in the second experimental group produce unrecognizable song, then it can be concluded that the Bobolinks learn their song. If despite exposure to foreign songs, the Bobolink sings the song of its parents, it can be concluded that song is inherited.
WORK CITED:
Ammer, F. K., and Capp, M. S. 1999. Song versatility and social context in
the bobolink. The Condor, 101(3), 686-688.
Avery, M., and Oring, L. 1977. Song dialects in the bobolink (Dolichonyx
oryzivorus). The Condor 79(1): 113-118.
Bent, A. C. 1958. Life histories of north american blackbirds, orioles,
tanagers, and allies. U.S. Natl. Mus. Bull. 211: 1-549.
Capp, M. 1992. Tests of the function of the song repertoire in bobolinks. The
Condor 94(2): 468-479.
Jacob, R.J., Steve, L.K., Sharon, A.K. 2016. Song adjustments by an open
habitat bird to anthropogenic noise, urban structure, and vegetation,
Behavioral Ecology 27(6): 1734–1744
Koper, N., Leston, L., Baker, T. M., Curry, C., and Rosa, P. 2016. Effects of
ambient noise on detectability and localization of avian songs and
tones by observers in grasslands. Ecology and Evolution 6(1):
245–255.
Wittenberger, J. 1983. A contextual analysis of two song variants in the
bobolink. The Condor 85: 172-184.
