

Image Courtesy of Wild About Birds
You’ve probably heard that dogs can only see in black and white, but that is just one of many examples of sensory differences between species. Like the visual system, the auditory system also differs dramatically between species as the result of different evolutionary pressures. The songbird auditory system is studied extensively as a model of animal communication because of songbirds’ complex vocal signaling. The black-capped chickadee (Poecile atricapillus) is a common songbird in the Northern United States, known for its distinctive chick-a-dee call. While information about the production and meaning of these vocalizations is readily available, not as much research has focused on how chickadees process sound.

A titmouse in one of our chickadee traps
Previous findings have provided a mechanistic overview of how sound is processed in black-capped chickadees (Knudsen & Gentner 2011). Auditory stimuli reach the birds’ inner ear where they resonate the basilar membrane of the cochlea. These vibrations are transduced into neural action potentials that are then carried to multiple cochlear nuclei in the brainstem. Neural pathways then lead out of the brainstem and head to a midbrain region called the mesencephalicus lateralis dorsalis (MLd). Afferent neurons project to auditory thalamic regions like the nucleus ovoidalis (Ov) and ultimately to areas of high interest to current researchers such as the caudomedial nidopallium (NCM) and the caudal mesopallium (CLM) (see Figure 1). However, few studies have related the physiology of their auditory system to their perception of sound in the wild.
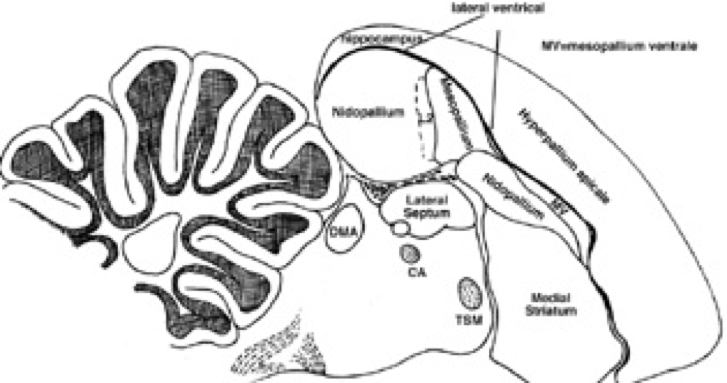
Figure 1. Hand drawn model of the central avian auditory system provided by C. Siang and E.D. Jarvis at Duke University. Many sensory neurobiologists have specifically focused their research on portions of the mesopallium and nidopallium
In the Sensory Ecology lab at Vassar College, we are exploring auditory processing in the black-capped chickadee using auditory brainstem responses (ABRs). Auditory brainstem responses are electrical signals generated by the auditory nerve and brainstem after the ear is stimulated by sound. They can be measured by inserting small needle electrodes beneath an anesthetized bird’s skin. When the bird is positioned directly beneath a speaker and sound clips are played, the electrodes record electrical activity in the neurons.
In this experiment, we played the chickadees two different types of sounds: broadband sounds and tonal sounds. Broadband sounds, like a drum beat or a clap, have energy across a wide band of frequencies while tonal sounds contain energy at a single frequency. A high-pitched treble and a low-pitched bass are examples of tonal sounds at different frequencies. Playing two different kinds of sounds allows us to ask several different questions about auditory processing in chickadees. By playing broadband noises and changing the rate at which we present them, we can gain information about how chickadees process sound at different speeds. Playing tonal sounds allows us to gain information about chickadees’ frequency sensitivity.
Frequency sensitivity can be assessed by playing tones of different frequencies at increasingly lower amplitudes until the chickadee’s neurons no longer produce a measurable response. The quietest sound that still produces a response at a given frequency is called the ABR threshold (see Figure 2).
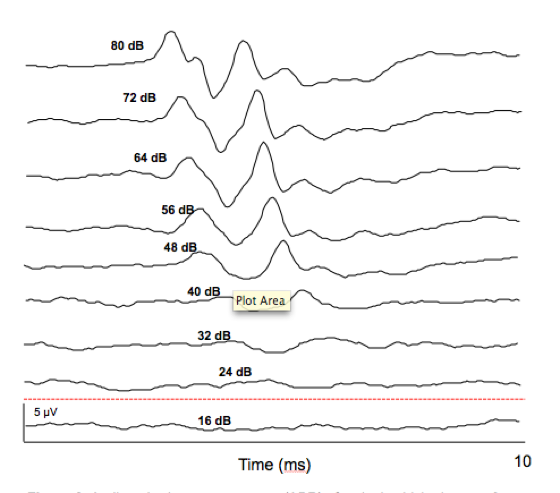
Figure 2. Auditory brainstem responses to a 3 kHz sound played at varying intensities, from loudest (80 dB) to softest (16 dB). The dotted red line at 20 dB indicates the ABR threshold at 3 kHz. From previous project by Wong & Gall (2015).
By finding the ABR threshold at a variety of frequencies, we can map out a chart of the chickadee’s auditory sensitivity. This chart, known as an audiogram, shows which ranges of frequencies chickadees can hear best (Figure 3). In one of our previous projects, we found that black-capped chickadees are most sensitive to sounds from 2-4 kHz, which, interestingly, corresponds to the major frequencies in their vocalizations. Determining frequency sensitivity and other aspects of chickadees’ auditory processing will allow us to make between-species comparisons and explore how different kinds of selection pressures may have influenced the evolution of the auditory system.
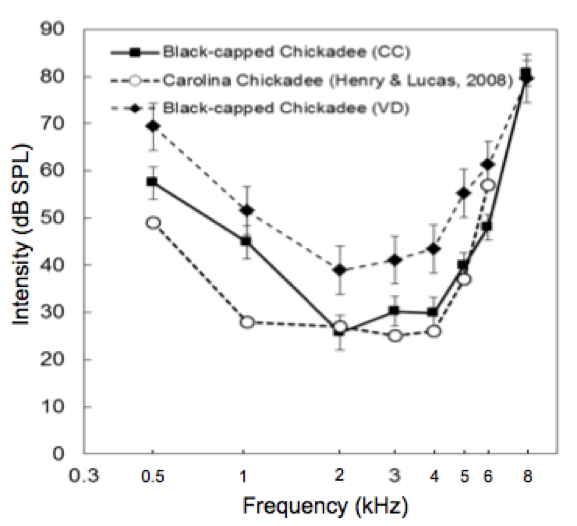
Figure 3. A chickadee audiogram from a previous project by Wong & Gall (2015). ABR thresholds are shown as a function of frequency. The black-capped chickadee is most sensitive to sounds from 2-4 kHz.
Auditory processing research is important to the wider field of animal behavior and physiology. Many researchers are interested in animal communication. They quantify different measurements of acoustic signals and examine the behavior of the receiving animal. However, humans view the world through our own senses and it can be easy to forget that animals may not be processing the same sensory stimuli. Without information about the acoustic abilities of the animals we study, we cannot completely understand how animals are sending and receiving the signals that are so important for everyday behavior. Chickadees, with their complex calls and songs, are popular study subjects. Therefore, understanding their ability to hear these calls and songs is extremely important to the field of animal communication.

Figure 4. (A) A chickadee found at Vassar Farm, (B) A spectrogram showing the frequency bands of a chickadee call, (C) A spectrogram of a chickadee song. From previous project by Wong & Gall (2015).
With our auditory research on frequency, we can study wild animals and how their behavior matches their hearing capabilities. We can answer questions like “Does the frequency range of chickadee signals fall inside of the range of the frequencies they can hear best?” and “How well can chickadees hear predator vocalizations?” We are also studying the ABRs of Northern saw-whet owls, tufted titmice, white-breasted nuthatches, gray treefrogs, and green treefrogs. Each species brings the possibility of new discoveries and a better understanding of the life on and above Vassar Farm’s forest floor.
References
Knudsen, D., & Gentner, T.Q. 2011. Mechanisms of Song Perception in Oscine Birds. Brain Language, 115: 59-68