
Social organisms experience a trade-off between the benefits of group living and the potential fitness costs generated by spatial crowding. The advantages of group living are partially offset by the cognitive challenges present in maintaining social boundaries. Group-living organisms typically navigate densely occupied social landscapes and minimize within-group conflict through the maintenance of social boundaries.

Carpenter Bee At Work (StockPhoto)
The sensory cues on which animals rely for nest localization vary with environmental conditions and with the sensory capabilities of the animal. The valley carpenter bee, Xylocopa varipuncta, nests in dead wood in large aggregations of up to several dozen nests that are only several centimeters apart. Previous research indicates that these bees emphasize the significance of visual cues, particularly landmark cues, but have also been shown to orient to specific chemical cues associated with nest entrances. Nest recognition mechanisms may integrate cues across multiple sensory modalities for a plethora of reasons. One popular hypothesis in current multimodality research states how the use of multiple modalities may provide ‘back-ups’ that either reinforce or conflict with the original message.

Female Carpenter Bee (Xylocopa varipuncta) (Wiki)
This study used female carpenter bees in order to investigate the importance of visual and olfactory cues in nest recognition ability. The distinguishing effects of proximal and distal visual landmarks in visual homing were studied as well as the effects of the presence and absence of olfactory cues in nests. Ostwald and colleagues hypothesized that both visual and olfactory cues facilitate nest localization.
The four treatment conditions assessed the effects of manipulated visual or olfactory cues at nest entrances. Methodology to examine each effect is summarized below.
- Distal Visual Cues –> Rotating the log (containing the nest) 180° about its vertical axis
- Proximal Visual Cues –> Moving colored shapes placed next to nest entrances
- Removal of Olfactory Cues à Entrances are rinsed with hexane
- Addition of Unfamiliar Conspecific Odors –> Application of cuticular hydrocarbon (CHC) extracts from nonresidents
All four treatment conditions measured search time and nest recognition error rate. Search time refers to when a returning bee begins entering a defined region and ends when the bee has entered her home nest. Nest recognition error rate refers to the number of nests a bee enters and exits until returning to her home nest.
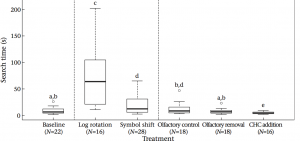
Figure 1. Time spent searching for the home nest during baseline, visual treatments (log rotation; symbol shift) and olfactory treatments (control; removal; cuticular hydrocarbon (CHC) extract). Letters indicate significant differences (P < 0.05); open circles represent outliers.
Search time was significantly increased in Treatment 1 with log rotation when compared to baseline. Foragers searched for approximately 8x longer and 4 out of the 16 bees inaccurately identified their home nest. A significant increase in search time was also present in Treatment 2 where geometric symbols were altered outside nest entrances. The same number of errors were made in this treatment when compared to the baseline control trials. Treatment 3, which removed chemical cues near the nest entrance, did not have a significant effect on search time. Two nest recognition errors were observed for both the control and manipulated trials. The additional of CHC from conspecifics in Treatment 4 resulted in a significant decrease in search time and interestingly, no errors were observed.
The word, significant, can be used to describe a majority of the results but the overarching question of what these comparisons mean has thus far been left unanswered. This nest recognition experiment examines the use of multimodal cue processing and demonstrates how the absence of olfactory cues does not affect the time it takes for female carpenter bees to find their home nest. Ultimately, all the foragers were able to locate their nest under each of the four experimental conditions which suggests that the current nest recognition system is robust to disruptions of individual cues.
The results support their hypothesis but interestingly enough, also suggest that visual cues alone may be sufficient in nest recognition. Chemical cues provide important individual or colony specific information but can also be unreliable. One of the most interesting results of this study was present in Treatment 4. The addition of scent cues actually helped bees find their nests more quickly rather than hindering their search. This suggests that carpenter bees may not demonstrate sensitivity to nest scent levels and are able obtain a sufficient amount of information from generic bee odors to aid nest localization. This also aligns with the notion about how multiple modalities may provide backups and reinforce visual information.

Carpenter Bee Entering Home Nest (Orkin)
This study demonstrates the use of primarily visual cues and how potentially weighted organization of cues can allow for prioritization of cues in order of stability and reliability. Furthermore, it is imperative that the environment is also taken into account when interpreting the results. The primary utilization of visual cues may be due to how the wood-nesting substrate provides more visual heterogeneity than soil, thus making visual navigation more reliable. Though not investigated in the experimental trials, nest recognition ability also functions in predator avoidance. During experimental trials, this study indirectly observed how bird predation is a significant risk for bees hovering outside nest entrances, especially when multiple prey attempts were displayed.
Yay! The bees were able to find their own nest AND avoid bird predation but how does this study contribute to the bigger picture in the field of sensory ecology? Ostwald and colleagues demonstrate how olfactory cues influence a bee’s ability to find her own nest, but the absence of these cues can also be overcome through other sensory systems such as visual. This is an imperative finding in the study of multimodal cues as well as the prioritization and relationship in sensory integration. As humans, we do not often think about how our multimodality contributes to our perceptions of the environment but multimodality is imperative in animal communication and increases signal detectability, provides redundancy and conveys complex messages. Carpenter bee aggregations may also result in intense nest competition thus nest localizations may have social implications for the maintenance of high-density nesting.
Now all that’s left to wonder about is how the bees inside felt when their neighbor bee entered the house unannounced! Awkward!
Reference: Ostwald, M. M, Schaffer, Z., Pratt, S., Fewell, J., (2019). “Multimodal Cues Facilitate Nest Recognition in Carpenter Bee Aggregations.” Animal Behaviour, Academic Press. Volume 155. Pg 45-51.
