
The art of communication involves two aspects: the first being the propagation of the signal across a medium, and the second being the reception of the information within the signal being communicated. The receiver can use this information to construct its response. Therefore, sending and receiving socio-sexual signals are immensely important to the overall fitness of a species, as these signals provide valuable information in terms of mate quality and availability.
In order to best survive and reproduce, living organisms behave in a particular way in response to their highly dynamic environments. An important factor within these environments is the interaction between rival males within a specific species, which is significant because the detection of a rival male has important implications for reproductive success.
Signals are transmitted and received using multiple sensory systems. Many times, multiple sensory cues are simultaneously propagated in communication to increase the success of the reception of the signal and its subsequent response. Previous research examined the impact of multiple sensory cues in the interaction in rival males in Drosophila, revealing that males within the D. melanogaster species demonstrate changes in mating behaviors in response to the presence of rival males. Another study found that D. melanogaster males use a variety of sensory cues, including olfactory, tactile, and auditory signals, to detect rival males and mount an appropriate behavioral response to the presence of another male in its environment.
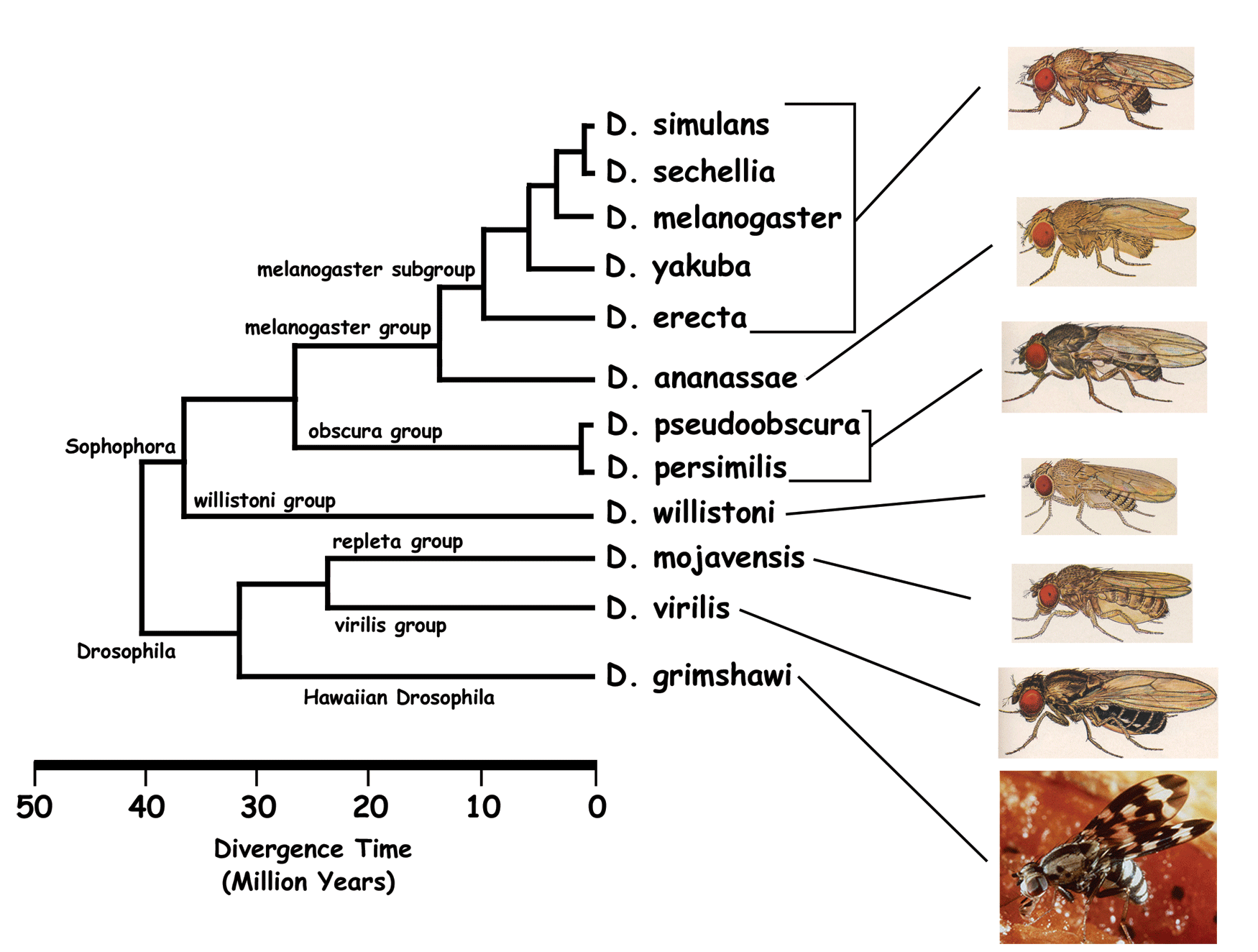
A phylogenetic tree of 12 Drosophila species, indicating the evolutionary relationship between D. melanogaster and D. pseudoobscura. (http://bioinformatica.upf.edu/2005/projectes05/3.2.1/Drosophila2.gif)
Maguire and colleagues (2015) expand on previous research concerning the perception of sensory cues and subsequent sexual behavior response in D. melanogaster males by extending it to a related species, D. pseudoobscura. In their study, the researchers aimed to discover whether males within the D. pseudoobscura species use a similar method of identification of rival males and demonstrate the same plasticity in terms of behavioral responses. They predicted that the studied males would use similar mechanisms due to the evolutionary relationship between the two species.
Wild females were collected and inbred to create lines that represented the genetic variation within the wild population of D. pseudoobscura. Males were collected from the standard line that Maguire et al. (2015) bred and randomly distributed into one of the 26 total treatments. Four sensory cues were tested: visual, auditory, olfactory, and tactile. Treatment groups were created for each sense, with the elimination of one sense in each condition, and then for the elimination of multiple senses in conditions to cover each possibility of sensory signaling in experimental treatments. Various alterations in methodology or the use of surgical procedures removed the ability to communicate with the examined sensory cues. Males were exposed to rivals for a period of four days before exposure to female counterparts.

An image of the experimental organism, D. pseudoobscura. (http://blogs.scientificamerican.com/compound-eye/files/2013/07/pseudoobscura1.jpg)
Statistical tests were used to measure the differences between exposed and control males in terms of copulation latency and copulation duration for each experimental condition. Their results revealed that males exposed to their rivals copulated significantly longer. Specifically, they found that visual cues are not important for detection of rival males. Rather, the presence of both olfactory and tactile cues is necessary for adequate rival detection and the following response to increase copulation duration to overcome rival mating pressure. Interestingly though, combining an auditory loss with either olfactory or tactile cue loss was associated with an increase in duration of copulation as well. Additionally, the study revealed that the male copulation latency was not affected by the presence of a rival male. Males did not rely on auditory signals to detect their rivals. Instead, the results indicated that the latency time increased after rival male exposure whenever auditory and any other sensory cue was eliminated, producing an overall confusing result as to the importance of auditory signals in male behavioral activity.
Maguire et al. (2015) suggest that this seemingly illogical result may be caused by a variety of factors. Removal of wings in the auditory treatment may have increased detection of the studied male due to a change in its scent or a change in its behavior, leading it to rely heavily on physical attacks to make its presence known. Increased fatigue in males from interactions with their rivals may have influenced the extent of sexual interactions with females.
Maguire and colleagues (2015) are quick to note potential flaws in their experimental design. They caution that the manner in which they eliminated the sensory cues may have produced unexpected results that influenced the way in which the results were analyzed. However, their results demonstrate that D. pseudoobscura males and D. melanogaster males differ in terms of their reliance on specific sensory cues in rival male detection. Overall, their results provide a foundation for future work regarding sensory cues and resulting sexual behavioral adaptations in mating systems, as their study suggests that the use of sensory cues in rival male detection may be the result of evolutionary change.
Reference:
Maguire C.P., Lizé A., Price T.A.R. (2015) Assessment of Rival Males through the Use of Multiple Sensory Cues in the Fruitfly Drosophila pseudoobscura. PLoS ONE 10(4): e0123058. doi: 10.1371/journal.pone.0123058
